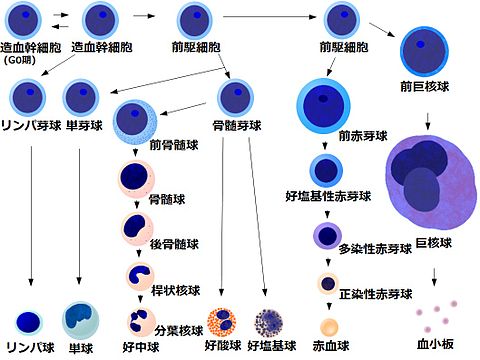
(Sertoli cell)
○セルトリ細胞とは
セルトリ細胞は、柱状の形態をとった精細管の上皮細胞である。セルトリ細胞間の密着結合によって血液精巣関門を形成し、また精子形成を助ける「ナース細胞」としての役割を持つ。1865年にこの細胞を発見したセルトリ博士からその名がつけられている。
関連:精子
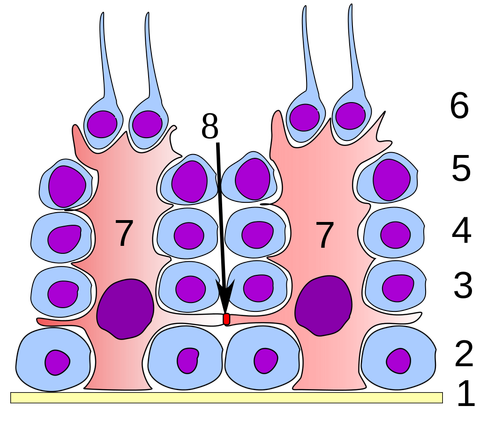
図1 セルトリ細胞
セルトリ細胞(7)は血液精巣関門(8)を形成し、生殖細胞の分化(1-6)を支えている。
○セルトリ細胞の形態
セルトリ細胞の形態学的な特徴は、①核が大きく、②Cytosolic Armと呼ばれる構造を持ち、③精細胞と接していることにある。セルトリ細胞の核は基底膜に近いところに位置し、球形ではなく刻みの入った形をしている。
クロマチン構造は密集していないユークロマチンが主であり、大きい核は薄くしか染色されないため、PAS染色での識別は困難である。その一方で、核内構造体である核小体は逆に非常に濃く染まり、近接する二つのChromosome center satelliteとともに、三つ一組で観察される。核を確認するためのマーカーとしては、AR(アンドロゲン受容体)やSOX9、WT1といった転写因子が用いられている。
Cytosolic Armは接触する精子を包み込んでいる、厚さわずか50nmの薄い構造体である。一つのセルトリ細胞から何本のもArmが伸びており、細胞の表面積は16000μm2にも及ぶ。直径10umの球形細胞の表面積が300μm2程度であるから、これは非常に大きな値であることがわかる。
○精子形成の補助
セルトリ細胞は精子分化のための環境を提供するとともに、各種シグナルによって分化を促進すると考えられている。精原細胞のみを体外に抽出したin vitroの条件でも精子は一定率で分化するが、その効率は精巣内部に大きく劣ることが確かめられている。
精原細胞から精細胞への分化過程にある細胞は、常にデスモソームやギャップ結合、接着結合を介してセルトリ細胞に接しながら、セルトリ細胞の細胞間隙を基底膜から内腔側へと移動する。内腔まで至った精細胞は鞭毛を内腔側に伸長していくにつれてセルトリ細胞の奥深くに貫入し、核の近くにまで迫る。セルトリ細胞の側ではApical Ectoplasmic specialization(ES) と呼ばれる構造が精細胞との接着面とアクチン繊維とをつなぎ留め、薄いArmの形態を支えている。
最終的に十分成熟した精細胞はアクチン―ミオシンモーターの力で内腔側へ押し返され、セルトリ細胞が精細胞の細胞質の大半を食作用によって回収し、精子は精細管内腔に放出される。放出された細胞質を残余体と呼ぶ。
セルトリ細胞はまた、内分泌されたFSH(卵胞刺激ホルモン)やテストステロンのシグナルを受容し、精細胞に転送する働きを持つ。シグナルを受容したセルトリ細胞はビタミンAを傍分泌し、精原細胞A型からA1型という段階への分化を促す。
○血液精巣関門
セルトリ細胞同士の密着結合とそれに結合するBasal ESは、血液精巣関門(Blood testis barrier : BTB)を形成する。血液精巣関門は血漿成分を完全に遮断するため、関門を超えて内腔側に移動した細胞は、すべてセルトリ細胞からの栄養供給を受ける。具体的には、例えば鉄イオン輸送に関して、セルトリ細胞がトランスフェリンを産生して鉄を集め、それを内腔側に放出していることが知られている。セルトリ細胞には精細胞の不要物を回収する働きもあり、グルタチオン還元酵素やCYP450といった薬物代謝酵素群の発現が顕著に高くなっている。
精原細胞からの分化過程での細胞移動の際には、セルトリ細胞は適切に密着結合やESの構造を組み替えて関門を乱さずに通過させることができる。一方で、血液精巣関門は免疫細胞や抗体を通過させないため、精細管の中は免疫の無い状態が維持され、免疫細胞による精子の攻撃が抑えられている。精子の形成が始まるのがすでに免疫寛容が終了した生誕後であるため、免疫細胞は精子を異物と認識してしまうのだ。睾丸の怪我で免疫細胞と精子が接触してしまうと精巣炎を発症し、最悪の場合男性不妊に陥るらしい。
セルトリ細胞は別個体に移植されても生存し、またともに移植された細胞をも免疫反応から守る働きがあるため、放出する物質によって化学的にも免疫阻害を行うと考えられている。具体的にはアポトーシス阻害物質(SERPINA3N)や補体阻害(CD55ほか)、抗炎症サイトカイン(TGFB1)などが報告されている。この機構をうまく利用して、糖尿病患者にインスリン放出β細胞とセルトリ細胞を移植するなどの医療応用が可能で
あろうと考えられている。
○幹細胞ニッチ
精巣において、幹細胞はセルトリ細胞と精細管基底膜との間の”幹細胞ニッチ”に所在しており、セルトリ細胞はGDNF(グリア細胞由来神経栄養因子)を放出することによってこの幹細胞ニッチを支えている。
GDNFは幹細胞の分化を抑制し、自己複製を促進する働きを持つ。GDNFの放出はFSHやFGF2による正の制御と、幹細胞が膜に生やしているNotchリガンド(DLL/JAG)の負の制御によって調節されている。幹細胞の増加はセルトリ細胞のNotchシグナルを活性化し、GDNFの放出抑制を引き起こす。幹細胞の減少はこの逆の効果を生み出すことで、幹細胞の数が維持されている。
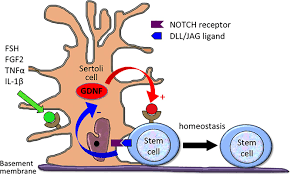
図2 幹細胞の維持
○参考文献
・The Sertoli Cell: one hundred fifty years of beauty and plasticity (L. R. Franca. et al., 2016)